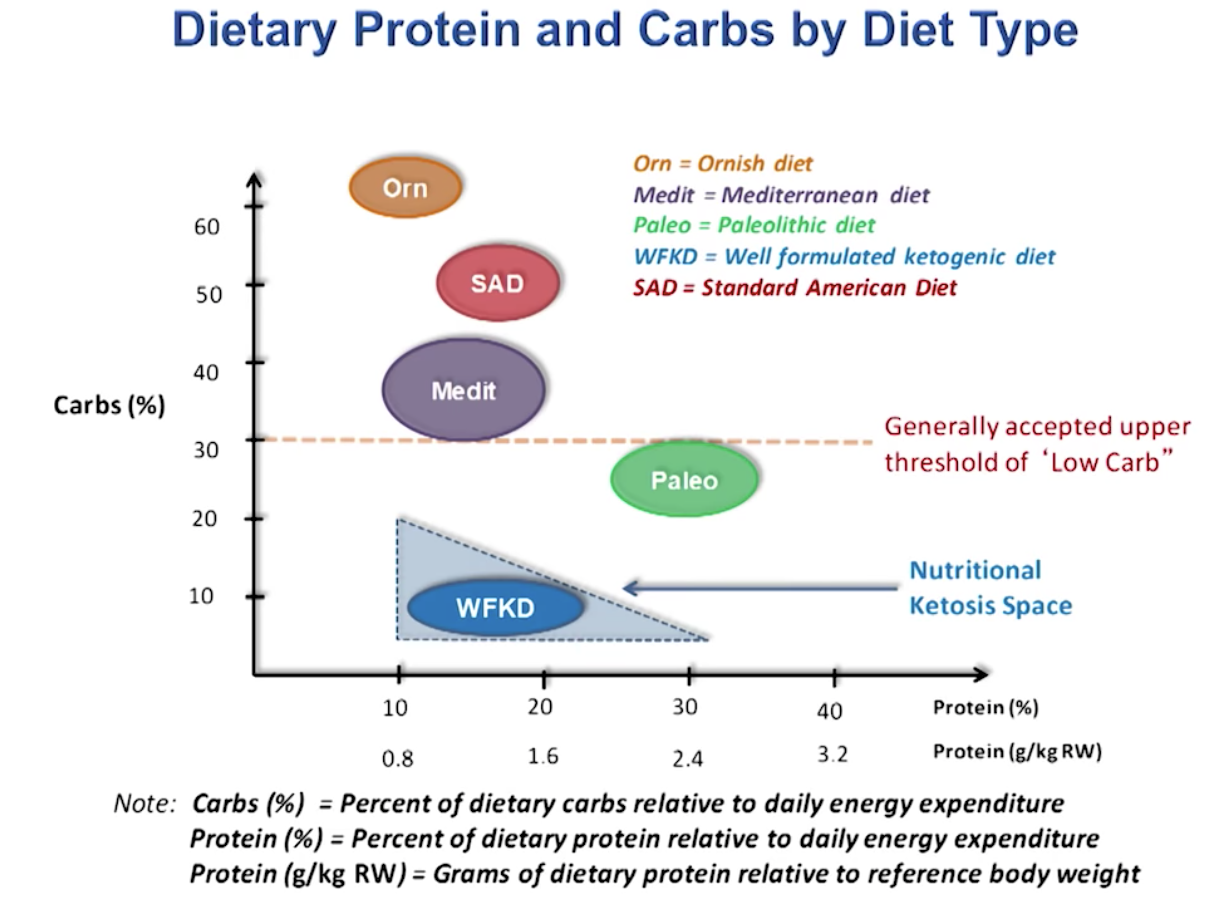
Ketogenic Diet
Highly anti inflammatory and anti cancer.
Best intervention to reduce insulin resistance.
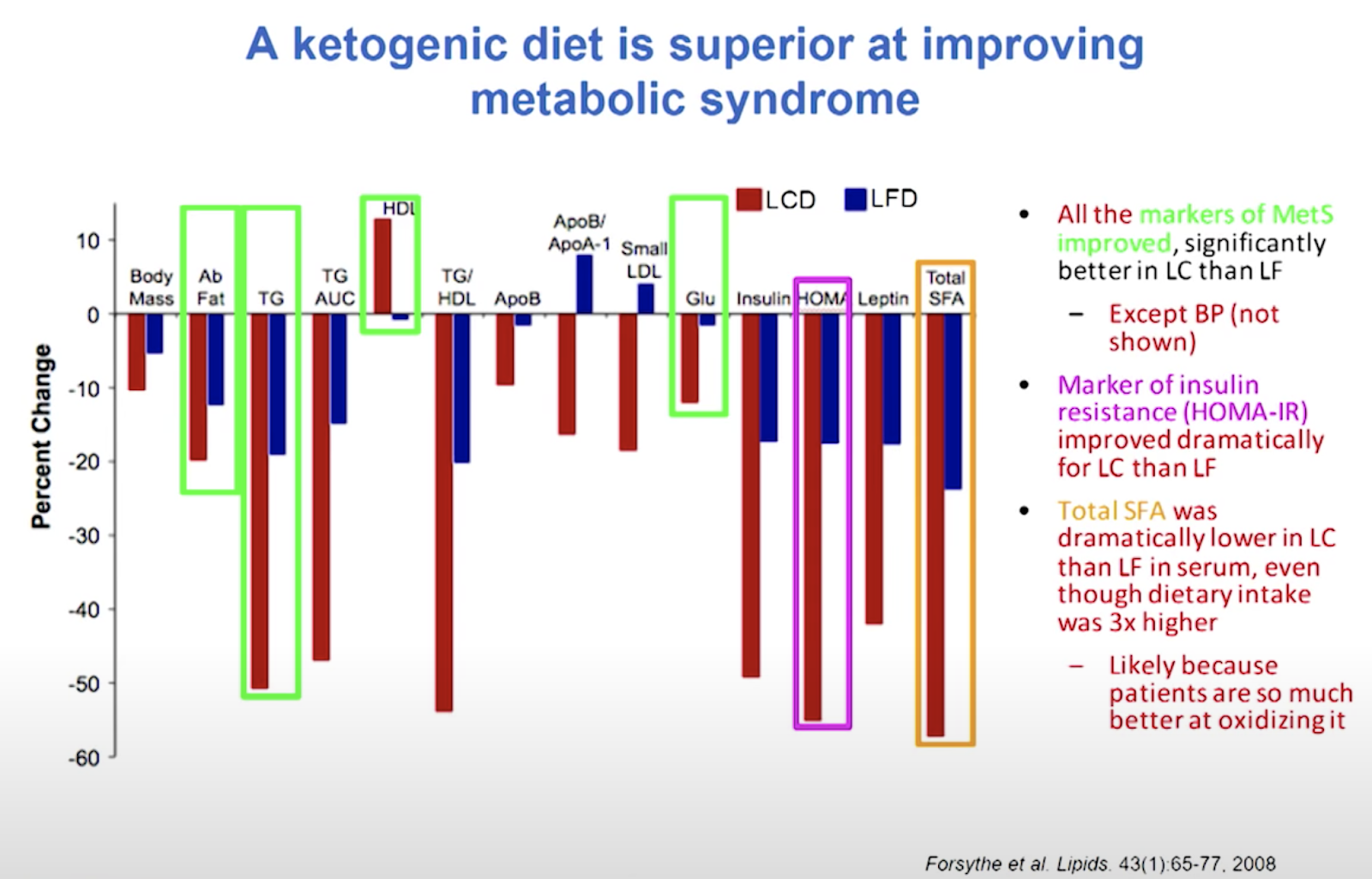
Human RCTs of 6-24 months duration gave fantastic results regarding controlling BP, insulin, and metabolic syndrome.
Anti-aging --- extended mice lifespan 10-13% --- think of it like an additional 10 yrs if you can do it all your life.
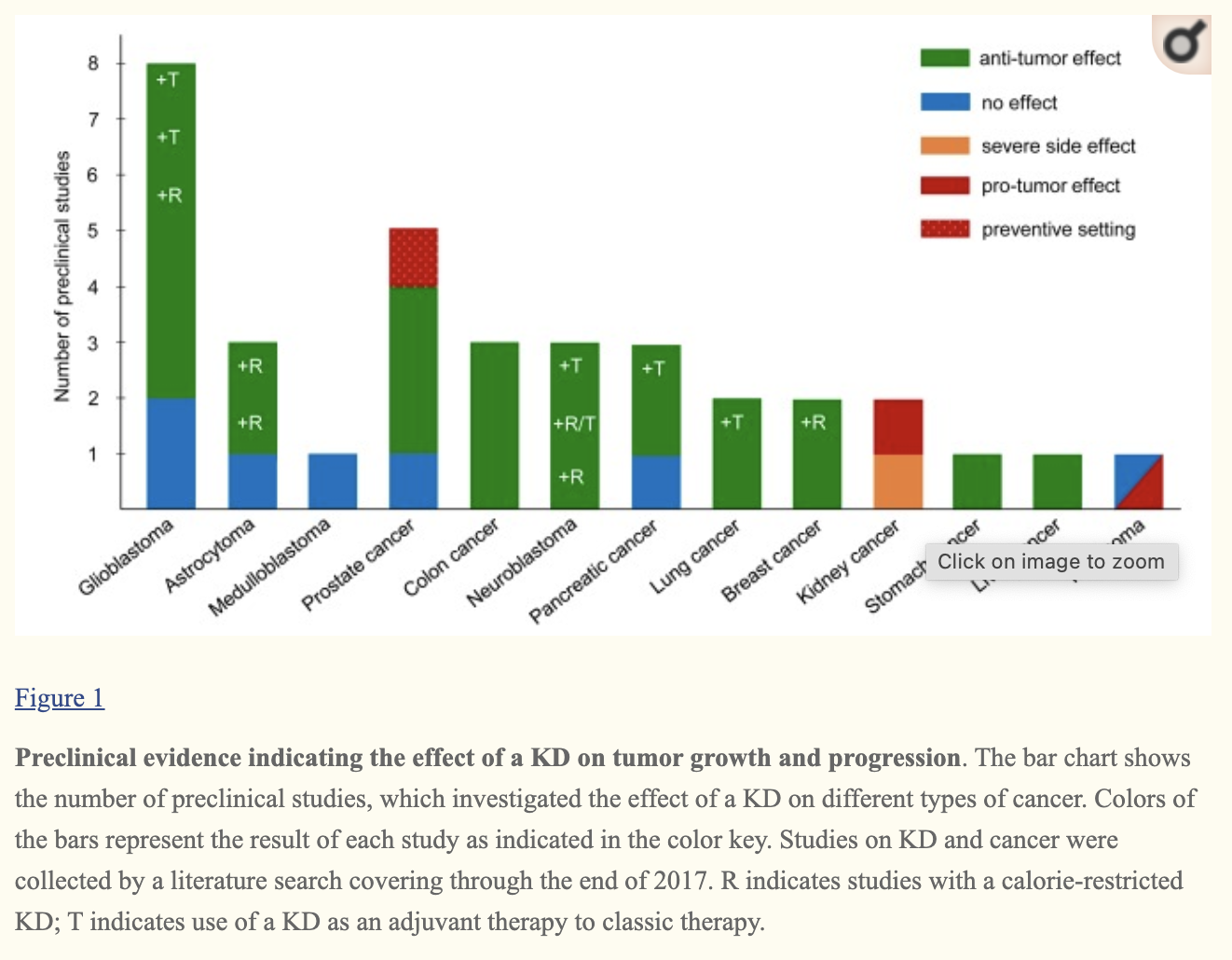
Regarding cancer, research on mice and small human trials showed it can (webmd, link):
Slow down tumor growth.
Help anti-cancer drugs work faster or better.
Ease inflammation, which can encourage cancer growth.
Protect healthy cells from damage from chemotherapy or radiation treatment.
Effect of KDs on proliferation is tumor type dependent (it doesn’t work for all cancers).
Low-carbohydrate and KDs increase quality of life of cancer patients.
Ketogenic diet in cancer therapy (2018)
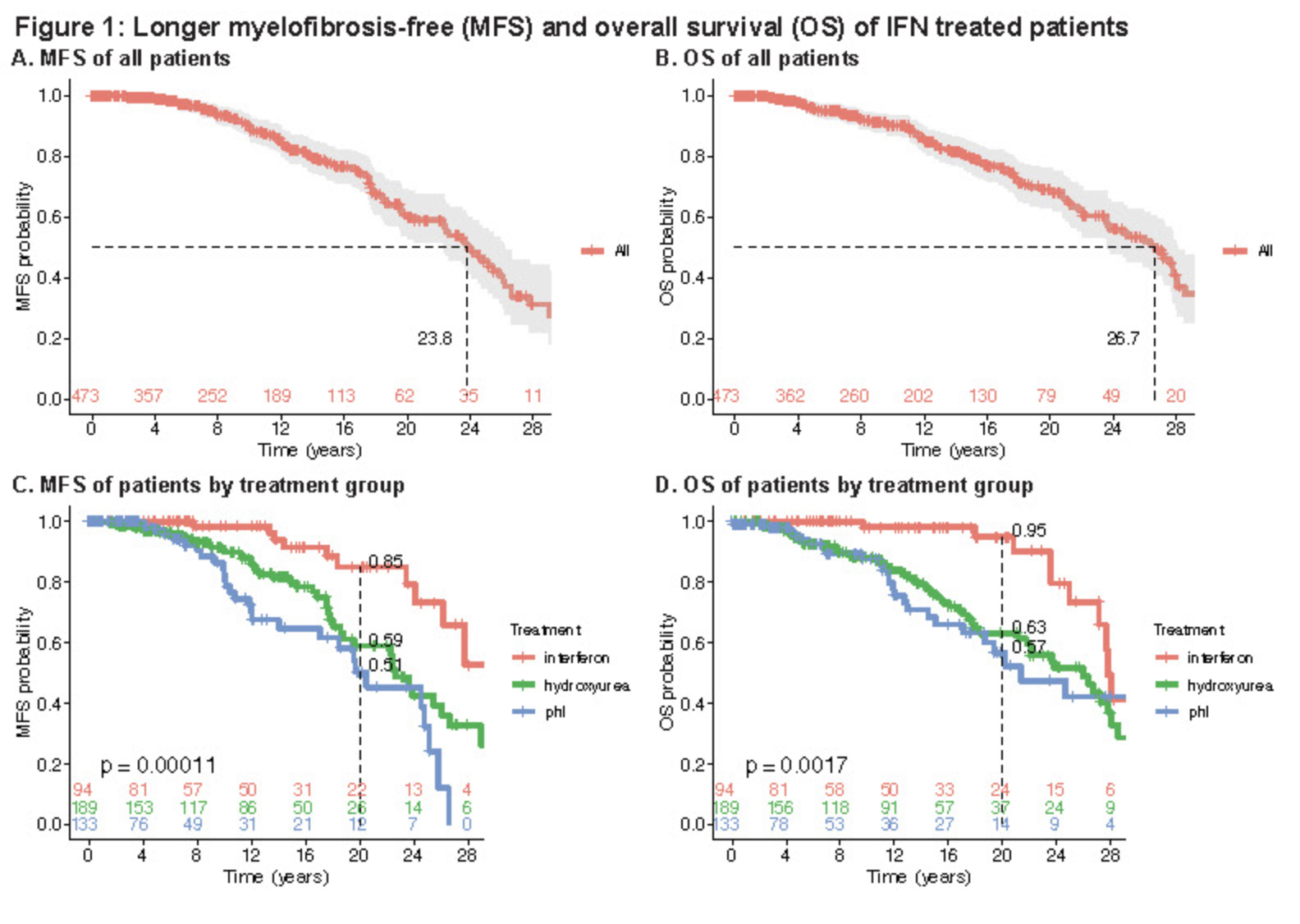
PV/MPN Testimonial
"regular phlebotomies (generally about every four weeks.) I started a diet plan that would put me in nutritional ketosis... I was able to go five months without a phlebotomy." (link)
Effect of Ketogenic Diet on the Clear Cell Renal Cell Carcinoma Cell Growth (link)
Mr. Benoit’s team tested a ketogenic diet (KD) on a ccRCC cell line in 15 mice. The team observed a significant decrease in tumor growth in the KD groups and, most remarkably, by the end of the 8-week study period, means tumor cell growth across the groups was 351% (Standard diet), 65% (Ketogenic diet 2:1), and 66% (Ketogenic diet 4:1), despite no difference in final weight.
[There’s a couple of negative publications on KD + Kidney Cancer, but if you read them they’re not high quality. ]
Dr. Eric Verdin on Ketogenic Diet Longevity, Beta-Hydroxybutyrate, HDAC Inhibitors & NAD+ (video)
Dr. John Ramsey - Ketogenic Diets and Aging (video)
Dr. Volek & Dr. Phinney - Translating the Basic Science of Nutritional Ketosis & Keto-Adaptation (video)
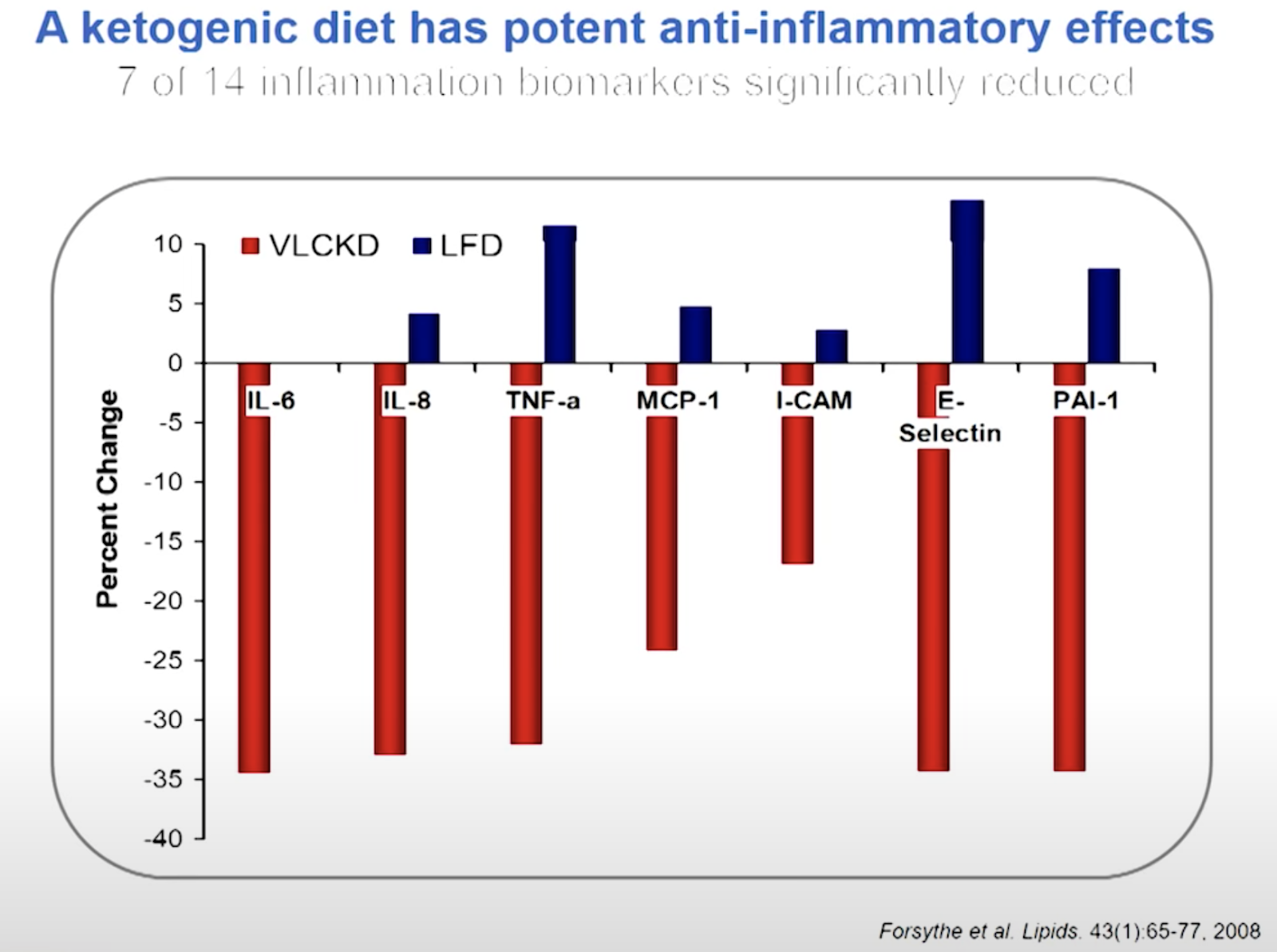
Steve Phinney - Inflammation, Nutritional Ketosis, and Metabolic Syndrome
https://www.youtube.com/watch?v=-ltCa513yZs
12 weeks ketogenic diet

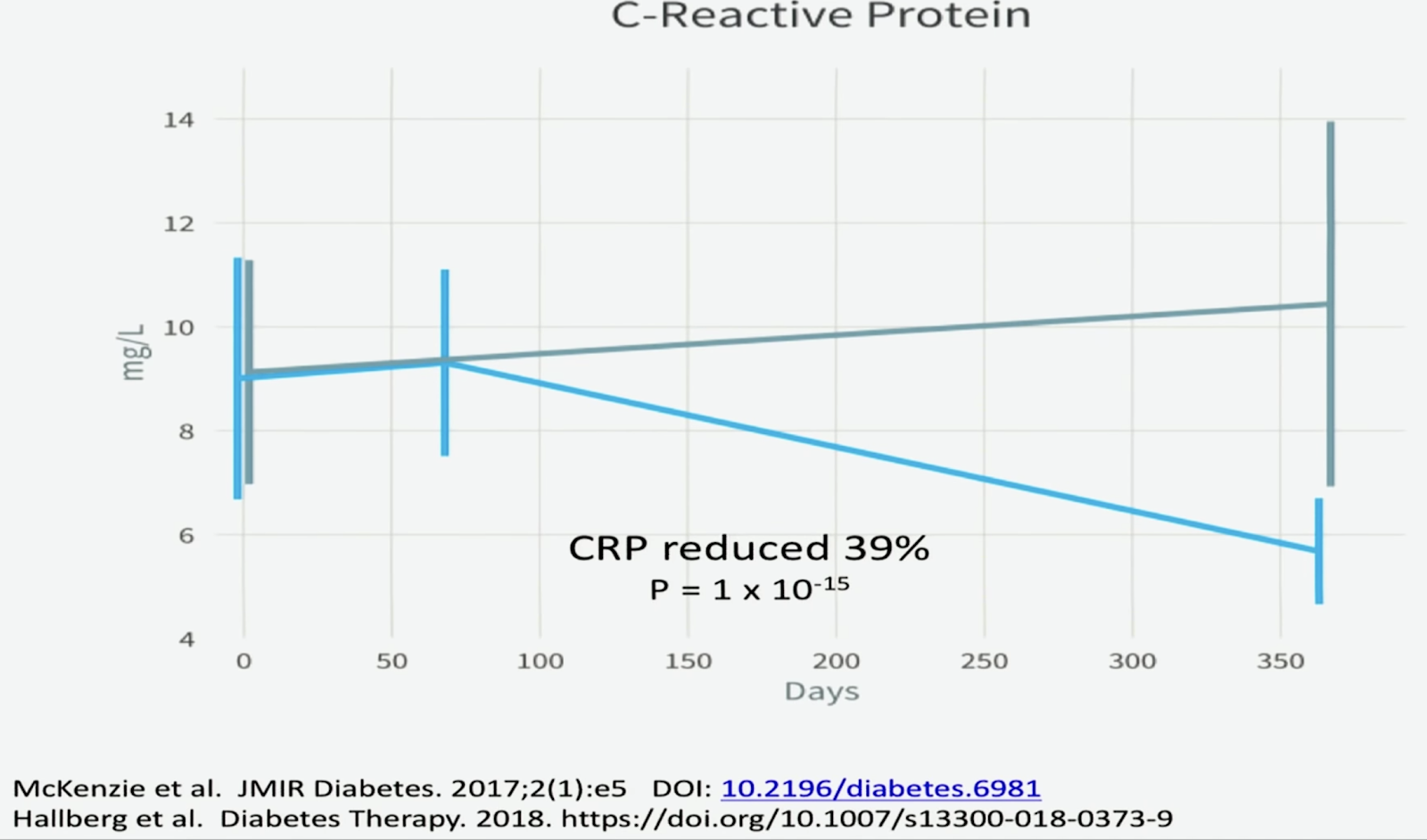

“Comparison of isocaloric very low carbohydrate/high saturated fat and high carbohydrate/low saturated fat diets on body composition and cardiovascular risk.” Nutrition & metabolism 3.1 (2006): 7. Noakes, Manny, et al.
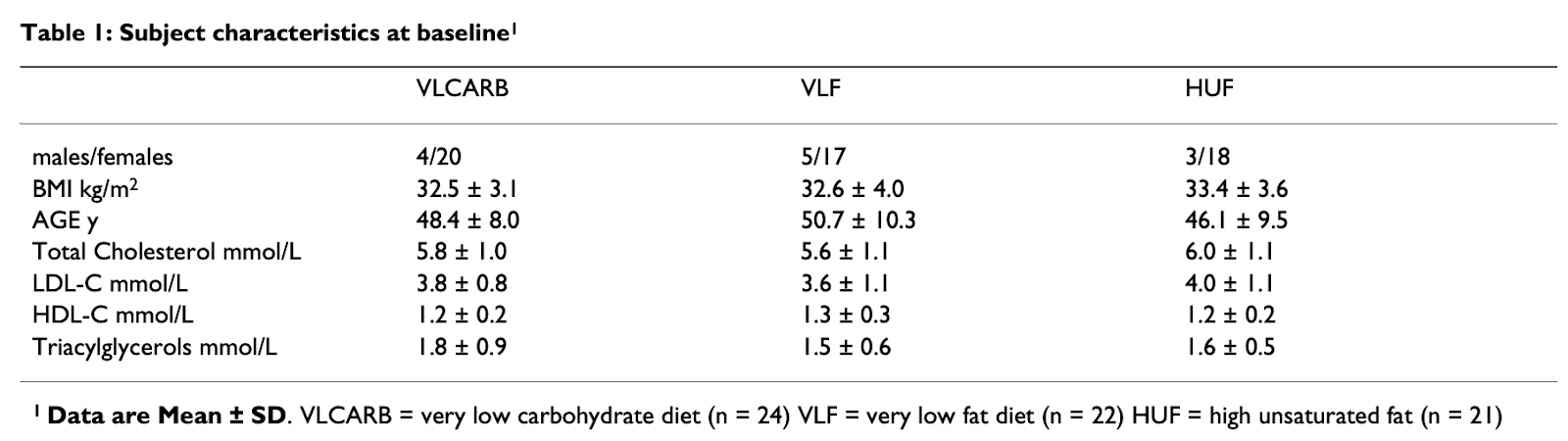
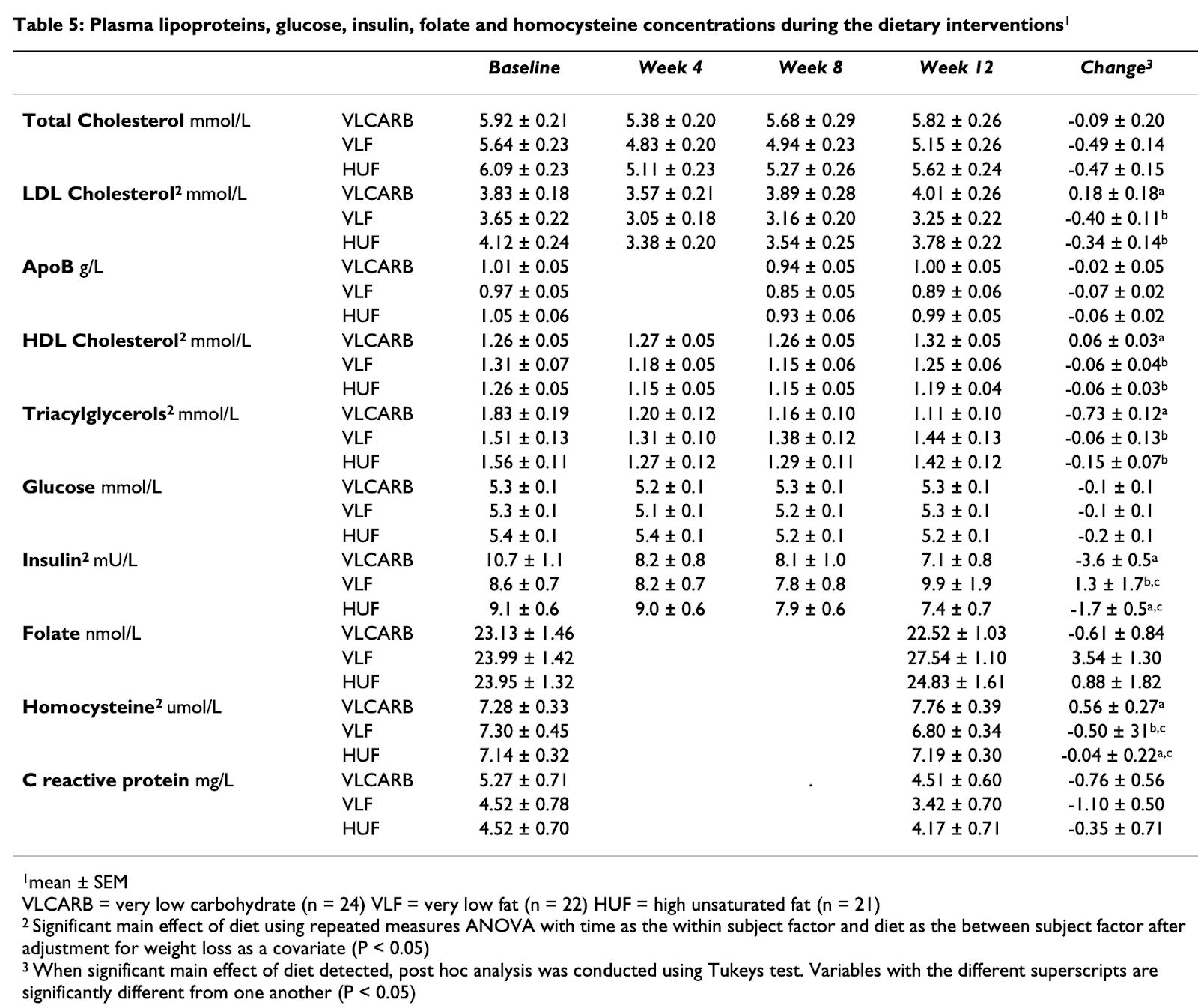
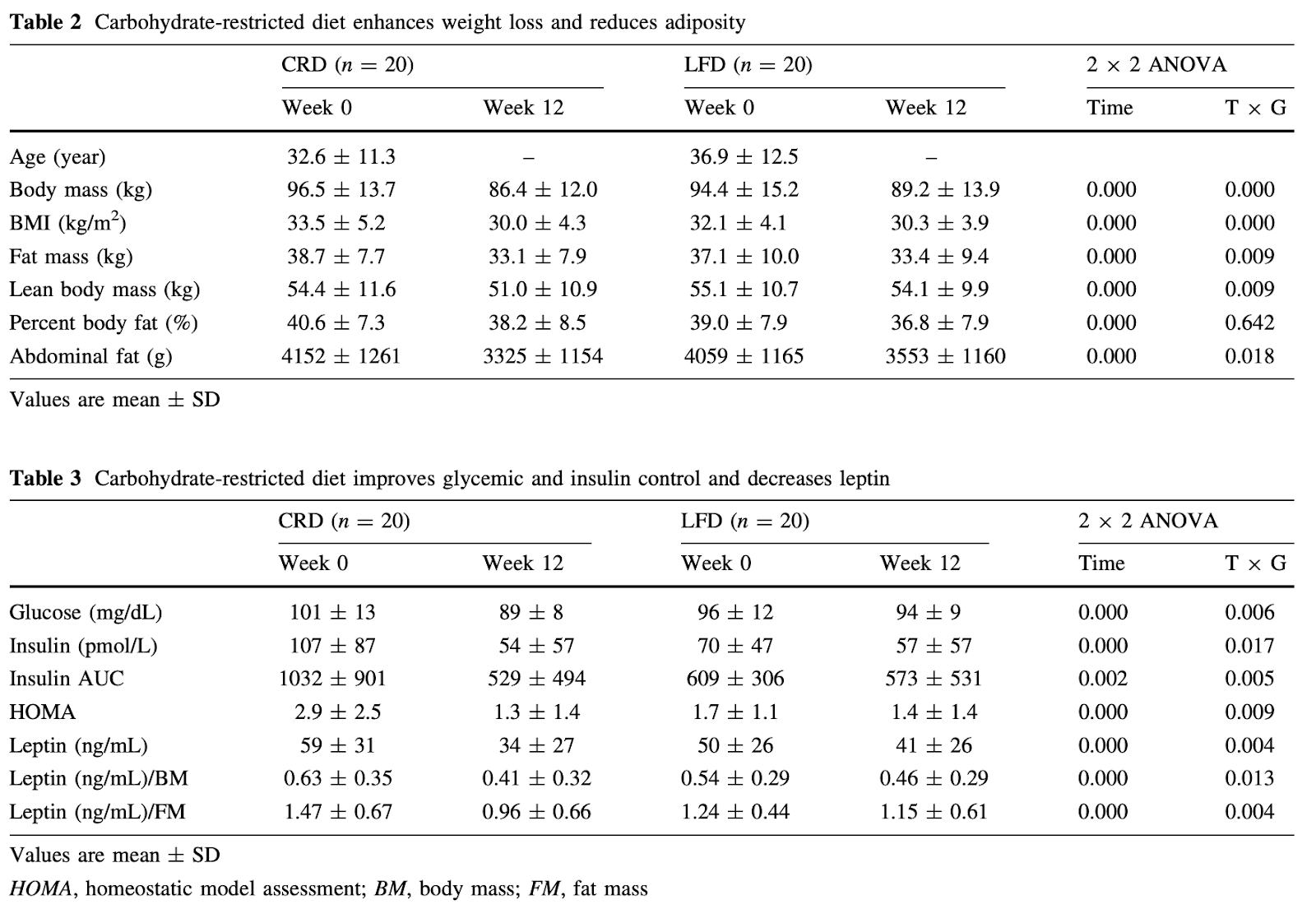
“Carbohydrate restriction has a more favorable impact on the metabolic syndrome than a low fat diet.” Lipids 44.4 (2009): 297-309. Volek, Jeff S., et al.
Insulin: 107 -> 15.4, 54 -> 7.7
Conversion: http://www.endmemo.com/medical/unitconvert/Insulin.php
A randomized trial of a low-carbohydrate diet vs orlistat plus a low-fat diet for weight loss. (2010)
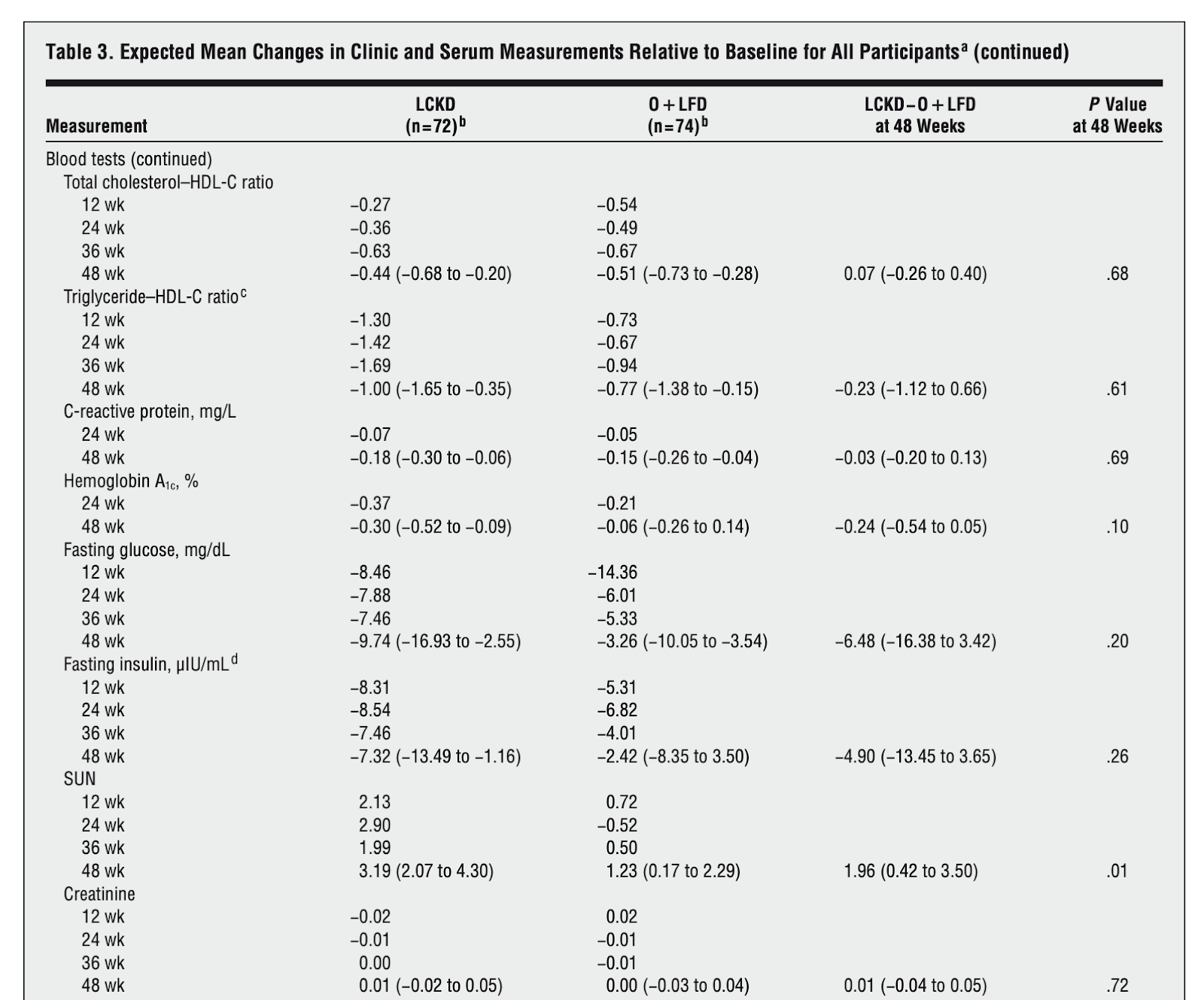
In 2010, researchers recruited 146 overweight and obese patients from the Department of Veterans Affairs in Durham, North Carolina. The mean age of these subjects was 52 years and the mean BMI was 39.3. Approximately 72% were men, 55% were African-American, and 32% had type 2 diabetes. The researchers placed the subjects in one of two weight-loss interventions that lasted 48 weeks.
The first group of 74 participated in a low-fat diet which had a 500-1000 kilocalorie/day deficit and less than 30% of calories from fat. Additionally, these subjects consumed three 120 mg tablets of the weight-loss orlistat drug per day. The second group of 72 ate a low-calorie ketogenic diet (LCKD) which initially had fewer than 20 grams of carbohydrates per day.
Overall, 52 out of 72 subjects in the LCKD group (79%) and 65 of the low-fat group (88%) completed the 48-week long trials. Both groups experienced similar levels of weight-loss and had similar levels of improvement in regards to heart-healthy high-density lipoprotein cholesterol (HDL-C) and triglycerides. However, only the LCKD group experienced improvements in levels of insulin, glucose, and hemoglobin A1C.
More specifically, the fasting insulin levels in the LCKD group decreased by 7.32 uIU/mL and 2.42 uIU/mL in the low-fat group. [8] Thus, fasting insulin levels in the LCKD subjects decreased by over three times compared to the low-fat subjects. Diabetes patients who completed the LCKD intervention experienced a 13.49 uIU/mL decrease in insulin while those that completed the low-fat intervention experienced an 8.35 uIU/mL decrease in insulin. [8] Collectively, these findings suggest that a LCKD diet has a stronger impact on insulin levels compared to low-fat diets supplements by drugs.
Associations of fats and carbohydrate intake with cardiovascular disease and mortality in 18 countries from five continents (PURE): a prospective cohort study
Background
The relationship between macronutrients and cardiovascular disease and mortality is controversial. Most available data are from European and North American populations where nutrition excess is more likely, so their applicability to other populations is unclear.
Methods
The Prospective Urban Rural Epidemiology (PURE) study is a large, epidemiological cohort study of individuals aged 35–70 years (enrolled between Jan 1, 2003, and March 31, 2013) in 18 countries with a median follow-up of 7·4 years (IQR 5·3–9·3). Dietary intake of 135 335 individuals was recorded using validated food frequency questionnaires. The primary outcomes were total mortality and major cardiovascular events (fatal cardiovascular disease, non-fatal myocardial infarction, stroke, and heart failure). Secondary outcomes were all myocardial infarctions, stroke, cardiovascular disease mortality, and non-cardiovascular disease mortality. Participants were categorised into quintiles of nutrient intake (carbohydrate, fats, and protein) based on percentage of energy provided by nutrients. We assessed the associations between consumption of carbohydrate, total fat, and each type of fat with cardiovascular disease and total mortality. We calculated hazard ratios (HRs) using a multivariable Cox frailty model with random intercepts to account for centre clustering.
Findings
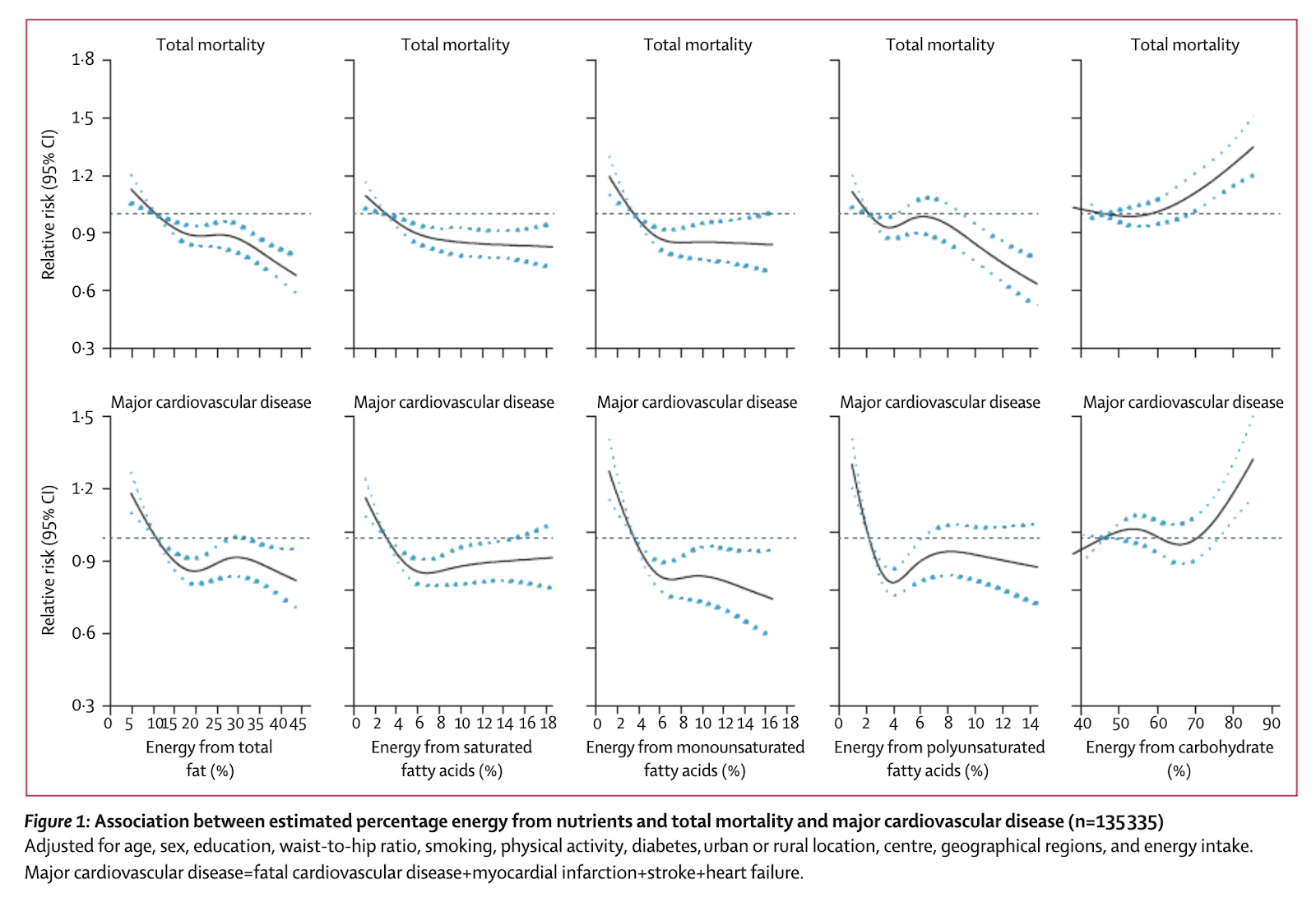
During follow-up, we documented 5796 deaths and 4784 major cardiovascular disease events. Higher carbohydrate intake was associated with an increased risk of total mortality (highest [quintile 5] vs lowest quintile [quintile 1] category, HR 1·28 [95% CI 1·12–1·46], p trend=0·0001) but not with the risk of cardiovascular disease or cardiovascular disease mortality. Intake of total fat and each type of fat was associated with lower risk of total mortality (quintile 5 vs quintile 1, total fat: HR 0·77 [95% CI 0·67–0·87], p trend<0·0001; saturated fat, HR 0·86 [0·76–0·99], p trend=0·0088; monounsaturated fat: HR 0·81 [0·71–0·92], p trend<0·0001; and polyunsaturated fat: HR 0·80 [0·71–0·89], p trend<0·0001). Higher saturated fat intake was associated with lower risk of stroke (quintile 5 vs quintile 1, HR 0·79 [95% CI 0·64–0·98], p trend=0·0498). Total fat and saturated and unsaturated fats were not significantly associated with risk of myocardial infarction or cardiovascular disease mortality.
Interpretation
High carbohydrate intake was associated with higher risk of total mortality, whereas total fat and individual types of fat were related to lower total mortality. Total fat and types of fat were not associated with cardiovascular disease, myocardial infarction, or cardiovascular disease mortality, whereas saturated fat had an inverse association with stroke. Global dietary guidelines should be reconsidered in light of these findings.
Since food patterns are culture dependent and dietary patterns are generally related to geographical region rather than income region, we categorised countries into seven regions. Regions included China, south Asia (Bangladesh, India, and Pakistan), North America, Europe (Canada, Poland, and Sweden), South America (Argentina, Brazil, Chile, and Colombia), Middle East (Iran, occupied Palestinian territory, Turkey, and United Arab Emirates), southeast Asia (Malaysia), and Africa (South Africa and Zimbabwe).
We calculated hazard ratios (HRs) using a multivariable Cox frailty model with random intercepts to account for centre clustering (which also adjusts for region and country).
In subgroup analyses, since higher carbohydrate (but lower fat) consumption is more common in Asian countries32,33 and lower carbohydrate intake (and higher fat) in non-Asian countries11 we examined whether the effect of carbohydrate and fats on outcomes were consistent in these two regions. The countries in Asia included Bangladesh, China, India, Malaysia, and Pakistan; the remaining countries were considered to be non-Asian countries.

Supplementation & Ketogenic Diet: A Winning Combination for Insulin? (link)
Provided the ketogenic diet’s positive impact on insulin resistance, can it be modified to confer even greater benefits?
In a study from earlier this year, researchers recruited 35 overweight subjects between 25 and 65 years old and divided them into two groups. The first group of 18 were instructed to follow a Mediterranean ketogenic diet for four weeks. They had an average age of 56.3 years and mean BMI of 29.34. Researchers instructed the second group of 17 subjects to follow the same ketogenic diet and also take three fish-oil supplements containing omega-3 fatty acids per day. These molecules are usually found in fatty fish, such as sardines and mackerel, and also in walnuts and chia seeds. These subjects had an average age of 58.1 years and a mean BMI of 29.17.
At the conclusion of the study, both groups showed a significant and similar decrease in BMI and per cent fat. Additionally, both showed a decrease in total cholesterol and low-density lipoprotein (LDL) but no significant change in HDL-cholesterol. These findings are in-line with many other research studies on the ketogenic diet.
The result for insulin proved particularly interesting. The subjects who consumed the ketogenic diet alone went from an average insulin concentration of 9.3 nIU/mL to an average insulin concentration of 7.4 nIU/mL of insulin. [11] This represents a 20.4% average decrease. In contrast, the subjects who consumed the ketogenic diet along with omega-3 fatty acid supplementation decreased their average insulin concentration from 11.1 nIU/mL to 6.3 nIU/mL. [11] This represents a 43% decrease in average insulin concentration- more than twice the decrease seen in the ketogenic diet alone. Additionally, subjects who took omega-3 fatty acid supplements experienced greater decreases in triglycerides and inflammation markers such as adiponectin and interleukin 1 beta (IL-1B). [11]
Supplementation & Ketogenic Diet: A Winning Combination for Insulin?
As a result of their findings, “the authors concluded that [omega-3] supplementation improved the positive effects of a ketogenic Mediterranean diet with phytoextracts on some cardiovascular/metabolic risk factors and inflammatory state.” [11]
Key Takeaways: Taking supplements of omega-3 fatty acids while on the ketogenic diet may confer additional benefits on metrics related to diabetes such as insulin resistance and levels of blood triglycerides and inflammatory markers in healthy, overweight subjects. However, these supplements do not affect other metrics related to diabetes such as fasting glucose.
Recommendations: If you have high levels of insulin, inflammation, or triglycerides are high, consider adding regular omega-3 fatty acid supplements (such as fish oil capsules) to your ketogenic diet.
Experiment with Supplements that Boost Insulin Sensitivity (link)
There are many supplements that can help with insulin sensitivity, but let’s stick with the ones that are backed by research:
Resveratrol. This is a polyphenolic compound that can be found in red wine and is known for its antioxidant benefits. Regarding insulin sensitivity directly, high-quality evidence indicates that resveratrol can boost glucose uptake as well.
Alpha Lipoic Acid. Alpha Lipoic Acid (ALA) is an organosulfur compound that is essential for aerobic metabolism. Many studies have reported that supplementation with this compound helps to reduce insulin resistance in subjects with type 2 diabetes.
Berberine. This is a plant alkaloid that has been shown to lower blood glucose in many cases. Some researchers have even found berberine to be as effective as the anti-diabetic medication metformin.
Chromium. Some evidence indicates that this essential trace element has the ability to enhance the effects of insulin and decrease blood glucose levels.
Magnesium. This essential mineral is so crucial for the effectiveness of insulin that magnesium deficiency can worsen insulin sensitivity.
Ketone bodies as signaling metabolites (link)
Traditionally, the ketone body β-hydroxybutyrate (βOHB) has been looked upon as a carrier of energy from liver to peripheral tissues during fasting or exercise. However, βOHB also signals via extracellular receptors and acts as an endogenous inhibitor of histone deacetylases (HDACs). These recent findings support a model in which βOHB functions to link the environment, in this case the diet, and gene expression via chromatin modifications. Here, we review the regulation and functions of ketone bodies, the relationship between ketone bodies and calorie restriction, and the implications of HDAC inhibition by the ketone body βOHB in the modulation of metabolism, and diseases of aging.
Ketogenic Metabolism Inhibits Histone Deacetylase (HDAC) and Reduces Oxidative Stress After Spinal Cord Injury in Rats
The aim of this study is to investigate the effect of ketogenic metabolism, induced by different diet interventions, on histone acetylation and its potential antioxidant capacity to injured spinal cord tissue in rats. 72 male Sprague-Dawley rats were randomly divided into 4 groups, fed with ketogenic diet (KD), every other day fasting (EODF), every other day ketogenic diet (EODKD) and standard diet (SD) respectively for 2 weeks. β-Hydroxybutyrate (βOHB) concentration was measured both in serum and cerebrospinal fluid (CSF). C5 spinal cord tissue was harvested before, at 3 h and 24 h after injury for analysis of HDAC activity, histone acetylation and oxidative makers. All three dietary interventions resulted in a significant increase of βOHB level in both serum and CSF, and inhibited HDAC activity by 31-43% in spinal cord. Moreover, the expressions of acetylated histone AcH3K9 and AcH3K14 were significantly increased. Anti-oxidative stress genes Foxo3a and Mt2 and related proteins, such as mitochondrial superoxide dismutase (SOD), FOXO3a, catalase were increased in dietary intervention groups. After SCI, high ketogenic metabolism demonstrated significant reduction of the expression of lipid peroxidation factors malondialdehyde (MDA), and this might contribute to the reported neuroprotection of the spinal cord from oxidative damage possibly mediated by increasing SOD. The result of this study suggested that by inhibiting HDAC activity and modifying related gene transcription, ketogenic metabolism, induced by KD, EODF or EODKD, might reduce oxidative damage in the spinal cord tissue after acute injury.
Prominent action of butyrate over β-hydroxybutyrate as histone deacetylase inhibitor, transcriptional modulator and anti-inflammatory molecule, 2019
Butyrate and R-β-hydroxybutyrate are two related short chain fatty acids naturally found in mammals. Butyrate, produced by enteric butyric bacteria, is present at millimolar concentrations in the gastrointestinal tract and at lower levels in blood; R-β-hydroxybutyrate, the main ketone body, produced by the liver during fasting can reach millimolar concentrations in the circulation. Both molecules have been shown to be histone deacetylase (HDAC) inhibitors, and their administration has been associated to an improved metabolic profile and better cellular oxidative status, with butyrate inducing PGC1α and fatty acid oxidation and R-β-hydroxybutyrate upregulating oxidative stress resistance factors FOXO3A and MT2 in mouse kidney. Because of the chemical and functional similarity between the two molecules, we compared here their impact on multiple cell types, evaluating i) histone acetylation and hydroxybutyrylation levels by immunoblotting, ii) transcriptional regulation of metabolic and inflammatory genes by quantitative PCR and iii) cytokine secretion profiles using proteome profiling array analysis. We confirm that butyrate is a strong HDAC inhibitor, a characteristic we could not identify in R-β-hydroxybutyrate in vivo nor in vitro. Butyrate had an extensive impact on gene transcription in rat myotubes, upregulating PGC1α, CPT1b, mitochondrial sirtuins (SIRT3-5), and the mitochondrial anti-oxidative genes SOD2 and catalase. In endothelial cells, butyrate suppressed gene expression and LPS-induced secretion of several pro-inflammatory genes, while R-β-hydroxybutyrate acted as a slightly pro-inflammatory molecule. Our observations indicate that butyrate induces transcriptional changes to a higher extent than R-β-hydroxybutyrate in rat myotubes and endothelial cells, in keep with its HDAC inhibitory activity. Also, in contrast with previous reports, R-β-hydroxybutyrate, while inducing histone β-hydroxybutyrylation, did not display a readily detectable HDAC inhibitor activity and exerted a slight pro-inflammatory action on endothelial cells.
The ketogenic diet inhibits the mammalian target of rapamycin (mTOR) pathway, 2011
The ketogenic diet (KD) is an effective treatment for epilepsy, but its mechanisms of action are poorly understood. We investigated the hypothesis that KD inhibits mammalian target of rapamycin (mTOR) pathway signaling. The expression of pS6 and pAkt, markers of mTOR pathway activation, was reduced in hippocampus and liver of rats fed KD. In the kainate model of epilepsy, KD blocked the hippocampal pS6 elevation that occurs after status epilepticus. As mTOR signaling has been implicated in epileptogenesis, these results suggest that the KD may have anticonvulsant or antiepileptogenic actions via mTOR pathway inhibition.
Life span extension by glucose restriction is abrogated by methionine supplementation: Cross-talk between glucose and methionine and implication of methionine as a key regulator of life span
Caloric restriction (CR) is known to extend life span across species; however, the molecular mechanisms are not well understood. We investigate the mechanism by which glucose restriction (GR) extends yeast replicative life span, by combining ribosome profiling and RNA-seq with microfluidic-based single-cell analysis. We discovered a cross-talk between glucose sensing and the regulation of intracellular methionine: GR down-regulated the transcription and translation of methionine biosynthetic enzymes and transporters, leading to a decreased intracellular methionine concentration; external supplementation of methionine cancels the life span extension by GR. Furthermore, genetic perturbations that decrease methionine synthesis/uptake extend life span. These observations suggest that intracellular methionine mediates the life span effects of various nutrient and genetic perturbations, and that the glucose-methionine cross-talk is a general mechanism for coordinating the nutrient status and the translation/growth of a cell. Our work also implicates proteasome as a downstream effector of the life span extension by GR.
Low glycaemic diets alter lipid metabolism to influence tumour growth
Dietary interventions can change metabolite levels in the tumour microenvironment, which might then affect cancer cell metabolism to alter tumour growth1,2,3,4,5. Although caloric restriction (CR) and a ketogenic diet (KD) are often thought to limit tumour progression by lowering blood glucose and insulin levels6,7,8, we found that only CR inhibits the growth of select tumour allografts in mice, suggesting that other mechanisms contribute to tumour growth inhibition. A change in nutrient availability observed with CR, but not with KD, is lower lipid levels in the plasma and tumours. Upregulation of stearoyl-CoA desaturase (SCD), which synthesises monounsaturated fatty acids, is required for cancer cells to proliferate in a lipid-depleted environment, and CR also impairs tumour SCD activity to cause an imbalance between unsaturated and saturated fatty acids to slow tumour growth. Enforcing cancer cell SCD expression or raising circulating lipid levels through a higher-fat CR diet confers resistance to the effects of CR. By contrast, although KD also impairs tumour SCD activity, KD-driven increases in lipid availability maintain the unsaturated to saturated fatty acid ratios in tumours, and changing the KD fat composition to increase tumour saturated fatty acid levels cooperates with decreased tumour SCD activity to slow tumour growth. These data suggest that diet-induced mismatches between tumour fatty acid desaturation activity and the availability of specific fatty acid species determine whether low glycaemic diets impair tumour growth.
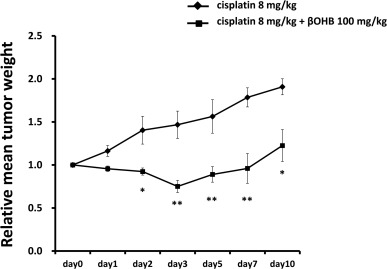
3.1. Efficacy of βOHB combined with cisplatin in HepG2 xenografts
First, we tested the combination of βOHB with cisplatin using HepG2 (an HCC cell line) xenografts to evaluate therapeutic efficacy in vivo. Tumor growth in the HepG2 xenograft was significantly suppressed by βOHB (100 mg/kg) + cisplatin (8 mg/kg) compared with cisplatin (8 mg/kg) alone (Fig. 1). No significant differences were observed in daily food intake or body weight changes between the cisplatin alone and βOHB + cisplatin groups (data not shown), and no deaths occurred in either group.
3.4. βOHB combined with cisplatin downregulated HDACs in HCC cells
D-β-Hydroxybutyric acid, along with butyric acid, are the two primary endogenous agonists of hydroxycarboxylic acid receptor 2 (HCA2), a Gi/o-coupled GPCR.[1][2][9]
β-Hydroxybutyric acid is able to cross the blood-brain-barrier into the central nervous system.[10] Levels of β-hydroxybutyric acid increase in the liver, heart, muscle, brain, and other tissues with exercise, calorie restriction, fasting, and ketogenic diets.[10] The compound has been found to act as a histone deacetylase (HDAC) inhibitor.[10] Through inhibition of the HDAC class I isoenzymes HDAC2 and HDAC3, β-hydroxybutyric acid has been found to increase brain-derived neurotrophic factor (BDNF) levels and TrkB signaling in the hippocampus.[10] Moreover, a rodent study found that prolonged exercise increases plasma β-hydroxybutyrate concentrations, which induces promoters of the BDNF gene in the hippocampus.[10] These findings may have clinical relevance in the treatment of depression, anxiety, and cognitive impairment.[10]
In epilepsy patients on the ketogenic diet, blood β-hydroxybutyrate levels correlate best with degree of seizure control. The threshold for optimal anticonvulsant effect appears to be approximately 4 mmol/L.[11]
Givinostat inhibits class I and class II histone deacetylases (HDACs) and several pro-inflammatory cytokines. This reduces expression of tumour necrosis factor (TNF), interleukin 1α and β, and interleukin 6.[8]
The Effect of Medium Chain Triglycerides on Time to Nutritional Ketosis and Symptoms of Keto-Induction in Healthy Adults: A Randomised Controlled Clinical Trial
In 28 healthy adults prescribed a ketogenic diet, randomised to receive either 30 ml of MCT, or sunflower oil as a control, three times per day, for 20 days.

the participants' BOHB levels were tested when fasted in the morning, and so, increased BOHB cannot be solely explained by transient ketonaemia resulting from MCT ingestion.

Suppression of Oxidative Stress by β-Hydroxybutyrate, an Endogenous Histone Deacetylase Inhibitor (link)
Concentrations of acetyl–coenzyme A and nicotinamide adenine dinucleotide (NAD+) affect histone acetylation and thereby couple cellular metabolic status and transcriptional regulation. We report that the ketone body d-β-hydroxybutyrate (βOHB) is an endogenous and specific inhibitor of class I histone deacetylases (HDACs). Administration of exogenous βOHB, or fasting or calorie restriction, two conditions associated with increased βOHB abundance, all increased global histone acetylation in mouse tissues. Inhibition of HDAC by βOHB was correlated with global changes in transcription, including that of the genes encoding oxidative stress resistance factors FOXO3A and MT2. Treatment of cells with βOHB increased histone acetylation at the Foxo3a and Mt2 promoters, and both genes were activated by selective depletion of HDAC1 and HDAC2. Consistent with increased FOXO3A and MT2 activity, treatment of mice with βOHB conferred substantial protection against oxidative stress.
Ketone supplementation decreases tumor cell viability and prolongs survival of mice with metastatic cancer, 2014
Cancer cells express an abnormal metabolism characterized by increased glucose consumption owing to genetic mutations and mitochondrial dysfunction. Previous studies indicate that unlike healthy tissues, cancer cells are unable to effectively use ketone bodies for energy. Furthermore, ketones inhibit the proliferation and viability of cultured tumor cells. As the Warburg effect is especially prominent in metastatic cells, we hypothesized that dietary ketone supplementation would inhibit metastatic cancer progression in vivo. Proliferation and viability were measured in the highly metastatic VM-M3 cells cultured in the presence and absence of β-hydroxybutyrate (βHB). Adult male inbred VM mice were implanted subcutaneously with firefly luciferase-tagged syngeneic VM-M3 cells. Mice were fed a standard diet supplemented with either 1,3-butanediol (BD) or a ketone ester (KE), which are metabolized to the ketone bodies βHB and acetoacetate. Tumor growth was monitored by in vivo bioluminescent imaging. Survival time, tumor growth rate, blood glucose, blood βHB and body weight were measured throughout the survival study. Ketone supplementation decreased proliferation and viability of the VM-M3 cells grown in vitro, even in the presence of high glucose. Dietary ketone supplementation with BD and KE prolonged survival in VM-M3 mice with systemic metastatic cancer by 51 and 69%, respectively (p < 0.05). Ketone administration elicited anticancer effects in vitro and in vivo independent of glucose levels or calorie restriction. The use of supplemental ketone precursors as a cancer treatment should be further investigated in animal models to determine potential for future clinical use.
Beta-hydroxybutyrate inhibits oncogenic signaling and cellular motility in pancreatic cancer cells
Pancreatic ductal adenocarcinoma is a devastating malignancy with a five-year survival rate less than 7%. It is characterized by a plethora of metabolic and signaling pathways, which plays a critical role in the pathogenesis of the disease. Caloric restriction and ketogenic diets have been shown to be beneficial in the management of several kinds of malignancies including pancreatic cancer. During caloric restriction or ketogenic diet supplementation, blood level of ketone bodies (beta-hydroxy butyrate or BHB, acetoacetate, acetone) gets elevated in the body. Among, these ketone bodies, BHB, the most abundant ketone body in mammals has been shown to act as the modulator of several signaling and inflammatory pathways. We have evaluated the effect of BHB on oncogenic signaling pathways and motility of pancreatic cancer cells. We observed reduced activation of key oncogenic signaling pathways, including mTOR. We also observed inhibition of cellular motility in a dose-dependent manner. Furthermore, we observed reduced expression of c-Myc, a key oncogene and a downstream effector molecule of mTOR. Correspondingly, we also observed reduced expression of several c-Myc target genes. We have also evaluated the effect of BHB on epithelial-mesenchymal transition (EMT) markers and observed altered expression of EMT markers like N-cadherin and E-cadherin. We also evaluated the effect of BHB on the expression of inflammatory cytokines in the pancreatic cancer cells and observed that BHB treatment leads to the reduction of expression of several inflammatory cytokines including IL6 and IL1-beta. Overall, our findings suggest that BHB inhibits inflammatory signaling in pancreatic cancer cells and reduces the cellular motility by modulating the expression of EMT regulators.
The action of β-hydroxybutyrate on the growth, metabolism and global histone H3 acetylation of spontaneous mouse mammary tumours: evidence of a β-hydroxybutyrate paradox
Ketone bodies have both metabolic and epigenetic roles in cancer. In several studies, they showed an anti-cancer effect via inhibition of histone deacetylases; however, other studies observed faster tumour growth. The related molecule butyrate also inhibits growth of some cancer cells and accelerates it in others. This “butyrate paradox” is thought to be due to butyrate mediating histone acetylation and thus inhibiting cell proliferation in cancers that preferentially utilise glucose (the Warburg effect); whereas in cells that oxidise butyrate as a fuel, it fails to reach inhibitory concentrations and can stimulate growth.
Methods
We treated transgenic mice bearing spontaneous MMTV-NEU-NT mammary tumours with the ketone body β-hydroxybutyrate (β-OHB) and monitored tumour growth, metabolite concentrations and histone acetylation. In a cell line derived from these tumours, we also measured uptake of β-OHB and glucose, and lactate production, in the absence and presence of β-OHB.
Results
β-OHB administration accelerated growth of MMTV-NEU-NT tumours, and their metabolic profile showed significant increases in ATP, glutamine, serine and choline-related metabolites. The β-OHB concentration within the treated tumours, 0.46 ± 0.05 μmol/g, had no effect on histone acetylation as shown by western blots. Cultured tumour cells incubated with 0.5 mM β-OHB showed β-OHB uptake that would be equivalent to 54% of glycolytic ATP phosphorylation and no significant change in glucose consumption or lactate production.
Conclusions
These results suggest that a β-OHB paradox may occur in these mammary tumours in a manner analogous to the butyrate paradox. At low β-OHB concentrations (<1 mM, as observed in our tumour model post-treatment), and in the absence of a Warburg effect, β-OHB is consumed and thus acts as an oxidative energy source and not as an epigenetic factor. This would explain the increase in tumour growth after treatment, the metabolic profiles and the absence of an effect on histone H3 acetylation.
Clinical effects of one year of chemotherapy with a modified medium-chain triglyceride ketogenic diet on the recurrence of stage IV colon cancer.
Background: Because many cancer cells display increased levels of glucose transporters that conduct glucose uptake and cancer cells use up to ten times more glucose than normal cells, glucide restrictions may be an effective nutritional therapy for patients with cancer. We introduced and examined effectiveness of modified medium-chain triglyceride (MCT) ketogenic diets with added MCT oil to suppress abdominal symptoms, as a severe carbohydrate intake restriction diet for patients with colon cancer. Methods: We administered chemotherapy in combination with the modified MCT ketogenic diet at a ratio of 1.4:1 for one year in 10 patients with stage IV recurrent colon cancer who experienced exacerbations under standard therapy (chemotherapy, best supportive care). Weight, albumin levels, presence of total blood ketone bodies, Response Evaluation Criteria in Solid Tumors (RECIST) classifications, and Quality of Life (QOL) evaluations were compared between patients in ketogenic diet group and 14 patients in the chemotherapy-alone group. Results: Although the patients’weight and serum albumin levels did not significantly differ in both groups, the serum total ketone bodies were significantly increased in the ketogenic diet group. The chemotherapy-alone group displayed a response rate of 21% and disease control rate of 64% across three Partial Response (PR), six Stable Disease (SD), and five Progressive Disease (PD) cases, but not in the Complete Response (CR) cases. The ketogenic diet group exhibited a response rate of 60% and disease control rate of 70% across five CR, one PR, one SD, and three PD cases. The combination of chemotherapy with the one year ketogenic diet increased the CR rates by 50%. Conclusions: Chemotherapy for one year in combination with a ketogenic diet presents higher response and disease control rates than chemotherapy alone. Therefore, the ketogenic diet may be a supportive therapy for patients with stage IV colon cancer.
Feasibility, Safety, and Beneficial Effects of MCT-Based Ketogenic Diet for Breast Cancer Treatment: A Randomized Controlled Trial Study, 2020
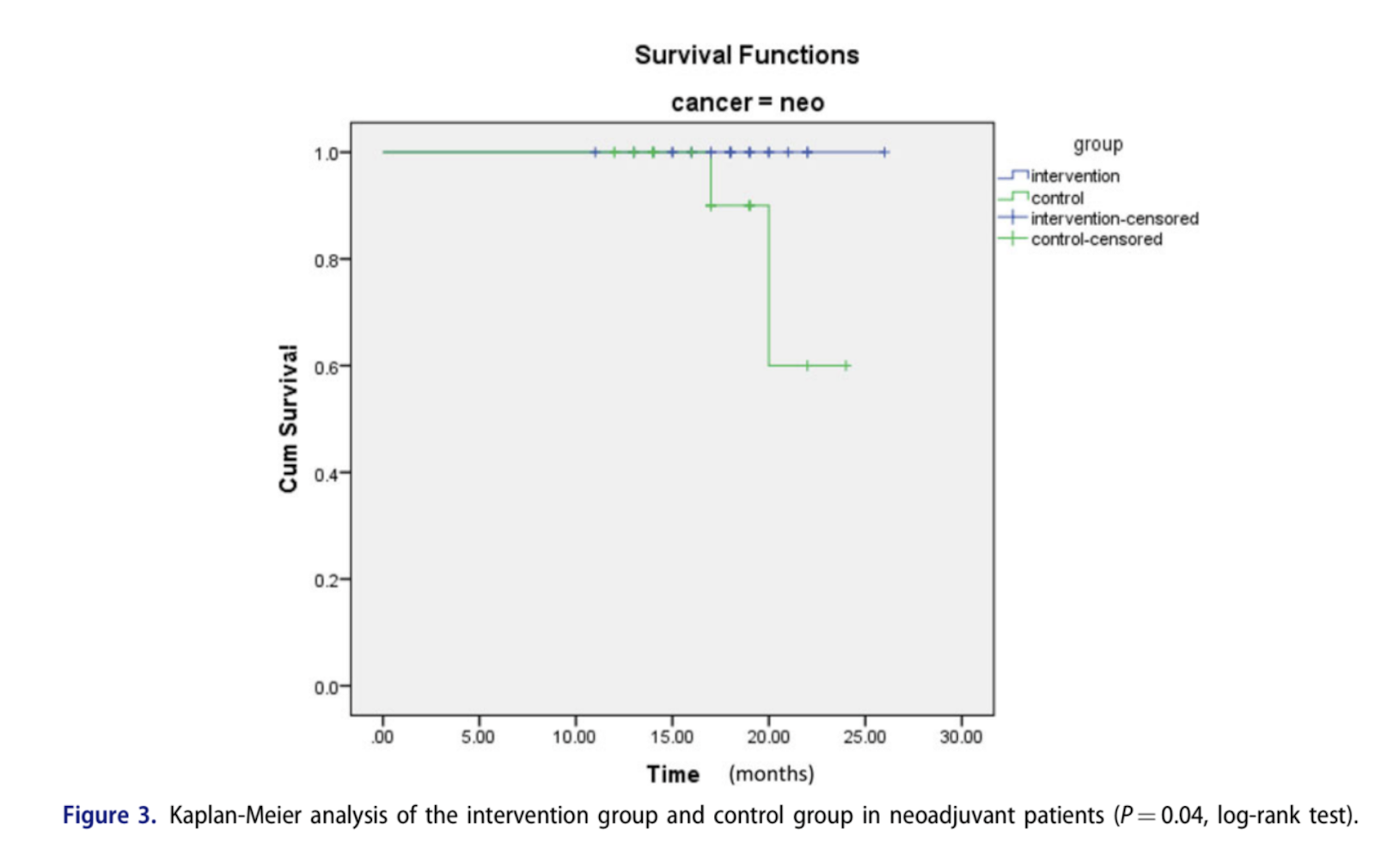
The present study was aimed to evaluate the safety, tolerability, and beneficial effects of a ketogenic diet (KD) on body composition and blood parameters and survival in patients with breast cancer. In this randomized, controlled trial, 60 patients with locally advanced or metastatic breast cancer and planned chemotherapy, were randomly assigned to a group receiving KDs (n = 30) or to a control group with standard diet (n = 30) for 3 months. Serum biochemical parameters and body composition were analyzed at baseline, every 3 weeks and end of each arm. Compliance and safety of KD were also checked weekly. Fasting blood sugar (FBS) was significantly decreased in intervention group compared to the baseline (84.5 ± 11.3 vs. 100.4 ± 11.8, P = 0.001). A significant inter-group difference was also observed for FBS level at end of intervention. There was an increasing trend in serum levels of ketone bodies in intervention group (0.007–0.92, P < 0.001). Compared to the control group, BMI, body weight, and fat% were significantly decreased in intervention group in last visit (P < 0.001). No severe adverse side effect was found regarding lipid profile and kidney or liver marker. Overall survival was higher in KD group compared to the control group in neoadjuvant patients (P = 0.04). Our results suggested that chemotherapy combined with KDs can improve the biochemical parameters, body composition, and overall survival with no substantial side effects in patients with breast cancer.
In the intervention group, a medium chain triglycerides (MCT)-based KD (contained 6% calorie from CHO, 19% protein, 20% MCT, 55% fat) was given to the patients for 90 consecutive days concurrently with the first 3 months of chemotherapy before surgical resection.
A volume of 500 ml of MCT oil from Nutricia Company were given to the patients every 2 weeks. For better tolerance MCT initiated with low dose and increased daily during 6 days until it reached the maximum dose and finally was discontinued in a stepwise process.

Clinical trial information: 000029527.
An intervention period: The glucide diet is three months by every meal.
A daily glucide intake: Less than 5% of intake energy.
Daily EPA(eicosapentaenoic acid) intake: More than 2 g.
A daily protein intake: More than weight * 1.6g.
A daily middle chain fatty acid intake: More than 40 g.

Comments